კორძიანი სხეული
კორძიანი სხეული, (/ˈkɔːrpəs kəˈloʊsəm/ ლათინურად „მკვრივი სხეული“) ასევე, კოჟრული კომისურა (შეერთება) — კომისურალური ნერვული ბოჭკოების ბრტყელი კონა, დაახლოებით 10 სმ სიგრძის, ცერებრალური თავის ტვინის ქერქის ქვეშ, ძუძუმწოვრების ტვინში. ის წარმოადგენს გასწვრივი ნაპრალის ნაწილს, ერთმანეთთან აკავშირებს მარცხენა და მარჯვენა ცერებრალურ ნახევარსფეროებს და ამ ნახევარსფეროებს შორის კომუნიკაციას უზრუნველყოფს. კორძიანი სხეული არის ყველაზე დიდი თეთრი ნივთიერების სტრუქტურის მქონე ერთეული ადამიანის ტვინში, შეიცავს რა 200-250 მილიონ აქსონურ პროექციას ან ბოჭკოს.[1]

სტრუქტურა რედაქტირება



კორძიანი სხეულის პოსტერიორულ დაბოლოებას, ნათხემის ახლოს, ეწოდება სპლენიუმი. ეს არის ყველაზე სქელი ნაწილი, რომელიც ნაწილობრივ ხურავს მესამე პარკუჭს, ტელა ქორიოიდეას და შუა ტვინს, და მთავრდება სქელ, ამოზნექილ, თავისუფალ ჩარჩოდ. სპლენიუმი ბერძნულად ითარგმნება როგორც სახვევი. ტვინის საგიტალური ჭრილი აჩვენებს, რომ კორძიანი სხეულის უკანა დაბოლოება ძლიერადაა წინ წახრილი ისე რომ, ზედა და ქვედა ნაწილები დართულია ერთმანეთზე.
კორძიანი სხეულის ანტერიორულ დაბოლოებას, შუბლის წილებთან ახლოს ეწოდება მუხლი. კორძიანი სხეულის მუხლი არის მოხრილი ქვემოთა და უკანა მიმართულებით გამჭვირვალე ტიხარის საპირისპიროდ და სწრაფად მცირდება სისქელეში. ის წაგრძელებულია უკან როგორც ტრიბუნა, (ამგვარად ეწოდება ჩიტის ნისკარტთან მსგავსების გამო) ქვემოდან უკავშირდება თხელ ტერმინალს, რომელიც გამოჭიმულია პარკუჭთაშორისი ღარიდან და ჩერდება ოპტიკური ღეროს ფუძესთან.
კორძიანი სხეულის არტერიული ტანი (ტრუნკუსი) არის „სახვევსა“ და „მუხლს“ შორის. შევიწროებულ ნაწილს ტანსა და „სახვევს“ შორის ეწოდება კორძიანი სხეულის ყელი.
კორძიანი სხეულის თითოეულ მხარეს ბოჭკოები წრიულად იხვევიან თეთრ ნივთიერებაში და მიემართებიან ცერებრალური ქერქის სხვადასხვა ნაწილებს, იხვევიან „მუხლიდან“ შუბლის წილის არტერიორულ საყრდენში, შემდეგ კი ისევ უკან ჩაიხვევიან კეფის წილში. ამ ორ ნაწილს შორის არის მოთავსებული ბოჭკოების მთავარი სხეული, რომელიც შეიცავს ტაპეტუმს, იგი იჭიმება ჰორიზონტალურად შუბლის წილის თითოეულ მხარეს და ფარავს გვერდითი პარკუჭის ცენტრალურ ნაწილს.
თხელი აქსონები კორძიანი სხეულის „მუხლში“, აკავშირებენ პრეფრონტალურ ქერქს ტვინის ორივე ნახევარს შორის, ეს ბოჭკოები ამოიზრდებიან ჩანგლის მსგავსი ბოჭკოების კონიდან, ტაპეტუმიდან და მიიმართებიან ანტერიორულად. უფრო სქელი აქსონები, შუა ტანში, ან კორძიანი სხეულის წვერო, აკავშირებს მოტორული ქერქის მიდამოებს. პროპორციულად დიდი ნაწილი კორძიანი სხეულისა მიძღვნილია დამატებითი მოტორული რეგიონებისთვის, რომლებიც მოიცავს ბროკას წილსაც. კორძის წინა სხეული, ცნობილი, როგორც „სახვევი“ (სპლენიუმი), კომუნიკაციას უწყობს ხელს და სომატოსენსორული ინფრომაციის გადაცემას უზრუნველყოფს შუბლის წილის ორ ნახევარს, ვიზუალურ ქერქსა და კეფის წილს შორის, მისი ბოჭკოები კი მიიმართებიან პოსტერიორულად .[2] [3] კვლევაში, რომელიც ჩატარდა ხუთიდან თვრამეტ წლამდე ასაკის ადამიანებზე, აღმოჩნდა დადებითი კორელაცია ასაკსა და კოჟრების სისქეს შორის.[1]
სქესობრივი დიმორფიზმი რედაქტირება
კორძიანი სხეული და მისი მიმართება სქესთან დებატის საგანი იყო სამეცნიერო და საერო სივრცეში საუკუნის მანძილზე. პირველმა კვლევამ ადრეულ მეოცე საუკუნეში აჩვენა, რომ კორძიანი სხეული განსხვავებული ზომის იყო ქალებსა და კაცებში. ეს კვლევა კითხვის ნიშნის ქვეშ დადგა და საბოლოოდ გზა დაუთმო უფრო მოწინავე „იმიჯინგის“ ტექნიკებს, რომელმაც წარმოაჩინა ადრეული კორელაციების სიმცდარე. თუმცა, გამოთვლითი ნეიროანატომიის მოწინავე ანალიტიკური ტექნიკები, განვითარდა 1990-იან წლებში, რომელთაც აჩვენეს, რომ სქესობრივი განსხვავებები იყო ნათელი, თუმცა შეიმჩნეოდა კორძიანი სხეულის მხოლოდ გარკვეულ ნაწილებში და ისინი კორელაციაში მოდიოდნენ კოგნიტური პერფორმანსის გარკვეულ ტესტებში. [4] ერთმა უახლესმა კვლევამ, რომელშიც იყენებდნენ მაგნიტურ რეზონანსულ ტომოგრაფიას აჩვენა, რომ შუა საგიტალური კორძიანი სხეულის განივი ნაწილი, ტვინის ზომის კონტროლის შედეგად, საშუალოდ პროპორციულად უფრო დიდი აღმოჩნდა ქალებში.[5]
დიფუზიური ტენზორული თანმიმდევრობების მაგნიტურ-რეზონანსულ მანქანებზე გამოყენებამ აჩვენა ზომა, რომლითაც მოლეკულების დიფუზია ხდება ქსოვილის შიგნით და გარეთ, სპეციფიკურ სივრცეში. ანიზოტროპია შეიძლება იყოს გათვლილი და გამოყენებული როგორც ანატომიური კავშირების სიმაგრის არაპირდაპირი გაზომვა. ამ თანმიმდევრობებმა აჩვენა შეთავსებადი სქესობრივი განსხვავებები ადამიანის კორძიანი სხეულის მორფოლოგიასა და მიკროსტრუქტურაში.[6] [7] [8] ასევე გამოყენებული იყო მორფომეტრული ანალიზი, რომ შეესწავლათ სპეციფიკური სამგანზომილებიანი მათემატიკური ურთიერთობები მაგნიტურ რეზონანსული ტომოგრაფიით, რამაც აჩვენა დამაჯერებელი და სტატისტიკურად მნიშვნელოვანი განსხვავებები სქესთა შორის.[9] [10] სპეციფიურმა ალგორითმება აჩვენა მნიშვნელოვანი განსხვავებები ორ სქესს შორის ერთ-ერთი კვლევის 70 % შემთხვევაში.[11]
სხვა კორელაციები რედაქტირება
ადამიანის კორძიანი სხეულის წინა ნაწილის შესახებ ნათქვამი იყო, რომ იგი მნიშნვნელოვნად დიდი იყო მუსიკოსებში ვიდრე არამუსიკოსებში,[12]ასევე იყო 0,75 სმ-ით [13] ან 11 პროცენტით დიდი ცაციებში და ადამიანებში, რომელთაც ორივე ხელის გამოყენება შეუძლიათ, ვიდრე მემარჯვენეებში.[13] [14] ეს განსხვავება აშკარაა კორძიანი სხეულის წინა და უკანა რეგიონებში, მაგრამ არა „სახვევში“.[13] სხვა მაგნიტურ-რეზონანსულმა მორფომეტრულმა კვლევამ აჩვენა, რომ კორძიანი სხეულის ზომა დადებით კორელაციაშია ვერბალური მეხსიერების ფუნქციონალურ აქტიურობასთან და სემანტიკური კოდირების ტესტის შესრულებასთან.[15] ბავშვებს, რომლებიც დაავადებულნი არიან დისლექსიით ტენდენციურად აქვთ პატარა და ნაკლებად განვითარებული კორძიანი სხეული, ვიდრე მათ ჯანმრთელ თანატოლებს.[16] [17] დადგინდა, რომ მუსიკალურმა ვარჯიშმა გაზარდა კორძიანი სხეულის პლასტიკურობა გარკვეული დროის სენსიტიური პერიოდის მსვლელობის განმავლობაში. ქვეტექსტი ამისა არის ხელების კოორდინაციის გაზრდა, განსხვავებები თეთრი ნივთიერების სტრუქტურაში და პლასტიკურობის გაზრდა მოტორულ და აუდიტორულ საშენში, რომელიც გამოიყენება შემდგომი მუსიკალური ვარჯიშის დასახმარებლად. კვლევამ შეისწავლა ბავშვები, რომლებმაც დაიწყეს მუსიკალური ვარჯიში ექვსი წლის ასაკამდე (მინიმუმ 15 თვიანი ვარჯიში), მათ აღენიშნებოდათ კორძიანი სხეულის მოცულობის ზრდა. ასევე მოზრდილებს, რომლებიც იწყებდნენ მუსიკალურ ვარჯიშს 11 წლის ასაკის შემდეგ, აღენიშნებოდათ ბიმანუალური კოორდინაციის ზრდა.[18]
კლინიკური მნიშვნელობა რედაქტირება
ეპილეფსია რედაქტირება

უკურნებელი ეპილეფსიის სიმპტომები შესაძლებელია იქნეს შემცირებული კორძიანი სხეულის ამოჭრით, ოპერაციისას, რომელსაც ეწოდება კორპუს კალოსოტომია.[19] ეს მეთოდი ჩვეულებრივად გამოიყენება შემთხვევებისთვის, რომელშიც კომპლექსური ან დიდი ეპილეფსიური შეტევები მიმდინარეობს ეპილოგენეტიკური ფოკუსით ტვინის ერთ მხარეს, ინტერჰემისფერული ელექტრული შტორმის გამოწვევით. შემზადება ამ პროცედურისთვის მოიცავს ელექტროენცეფალოგრამას, „MRI” “PET სკანირებას” და სპეციალიზებული ნევროლოგისტის, ნეიროქირურგის, ფსიქიატრის და ნეირორადიოლოგის შეფასებას, ვიდრე ოპერაცია ჩაითვლება საჭიროდ.[20]
შეფერხება განვითარებაში რედაქტირება
კორძიანი სხეულის ფორმირება იწყება პირველი შუალედური აქსონების გადაჯვარედინებით დაახლოებით ადამიანის განვითარების პრენატალური პერიოდის მე-12 კვირას,[21] ხოლო თაგვის ემბრიონული პერიოდის მე-15 დღეს.[22] კორძიანი სხეულის აგენეზისი არის იშვიათი კონგენიალური დაავადება, რომელიც არის ერთ-ერთი ყველაზე ჩვეულებრივი ტვინის მალფორმაცია, რაც შესწავლილა ადამიანებში და რომლის შემთხვევაშიც კორძიანი სხეული ნაწილობრივ ან სრულიად არ არსებობს.[23] კორძიანი სხეულის აგნეზიის დიაგნოსტიკა ჩვეულებრივ ხდება სიცოცხლის პირველი ორი წლის განმავლობაში და ის შესაძლოა ჩამოყალიბდეს, როგორც მწვავე სინდრომი პატარაობისას და ბავშვობისას, უფრო რბილი ახალგაზრდებსა და მოზრდილებში, ან მას საერთოდ არ ჰქონდეს სიმპტომები, იყოს ასიმპტომატური და შემთხვევით გამოვლენილი. კორძიანი სხეულის აგენეზის საწყისი სიმპტომები ჩვეულებრივ მოიცავს შეტევებს, რომლებსაც შეიძლება მოსდევდეს კვების პრობლემები და შეფერხებები თავის აღმართვაში, ჯდომაში, დგომაში და სიარულში. სხვა შესაძლო სისტემები შეიძლება მოიცავდეს გაუარესებებს მენტალურ და ფიზიკურ განვითარებაში, ხელისა და თვალის კოორდინაციაში, ვიზუალურ და აუდიტორულ მეხსიერებებში, ასევე შეიძლება განვითარდეს ჰიდროცეფალია. მსუბუქ შემთხვევებში სიმპტომები როგორიცაა შეტევები, განმეორებადი ლაპარაკი, ან თავისტკივილი შესაძლოა არ გამოვლინდეს წლების განმავლობაში. ზოგიერთი სინდრომი რომელიც ხშირად ასოცირდება კორძიანი სხეულის აგენეზთან არის აიკარდის სინდრომი, ანდერმანის სინდრომი, შაპიროს სინდრომი და აკროკალოსალური სინდრომი.
კორძიანი სხეულის აგენეზისი ჩვეულებრივ არ არის ფატალური. მკურნალობა ჩვეულებრივ მოიცავს სიმპტომების მენეჯმენტს, როგორებიცაა ჰიდროცეფალია და შეტევები, იმ შემთხვევაში თუ ისინი გამოვლინდებიან. ასევე, ამ დაავადების მქონე ბავშვების დიდ ნაწილს აქვს ნორმალური ცხოვრება და საშუალო ინტელექტი, მიუხედავად ამისა ყურადღებიანი ნეიროფსიქოლოგიური ტესტი ააშკარავებს ფაქიზ განსხვავებებს მაღალი ქერქის ფუნქციონირებაში, იგივე ასაკის და განათლების მქონე ინდივიდებში, რომელთაც არ აღენიშნებათ კორძიანი სხეულის აგენეზი. ბავშვები კორძიანი სხეულის აგენეზით, რომელთაც აღენიშნებათ ჩამორჩენა განვითარებაში ან შეტევითი აშლილობები,ასევე შემოწმებული უნდა იყვნენ მეტაბოლურ დაავადებებზეც.[24]
კორძიანი სხეულის აგენეზთან ერთად, მსგავსი სიმპტომები ახასიათებს ჰიპოგენეზს ( ნაწილობრივი ფორმირება), დიზგენეზისს, (მალფორმაცია) და ჰიპოპლაზიას (განვითარების დაბალი დონე, გადაჭარბებული სიგამხდრის ჩათვლით) უახლესმა კვლევებმა, ასევე, თვალსაჩინო გახადა შესაძლო კორელაციები კორძიანი სხეულის მალფორმაციასა და აუტიზმის სპექტრის დაავადებებს შორის.[25]
კიმ პიკი, დიდი მეცნიერი და ფილმის „წვიმის კაცი“ ინსპირაცია, ასევე დაავადებული იყო კორძიანი სხეულის აგენეზისით.
სხვა დაავადებები რედაქტირება
წინა კორძიანი სხეულის დაზიანებებმა შესაძლოა გამოიწვიოს აკინეტიკური მუტიზმი ან ანომიური აფაზია. იხილეთ ასევე:
- უცხო ხელის სინდრომი
- ალექსია აგრაფიის გარეშე (შემჩნეულია კორძიანი სხეულის „სახვევის“ დაზიანებისას)
- ნაპრალოვანი ტვინი
- სეპტო-ოპტიკური დისპლაზია (დე მორსიერის სინდრომი)
- გაფანტული სკლეროზი „დავსონის თითის“ ნიშნით
- ენცეფალოპათია, ზომიერი ფორმა კორძიანი სხეულის „სახვევის“ შექცევადი დაზიანებით- იშვიათი ენცეფალოპათია გაურკვეველი წარმოშობით, სახვევის გარდამავალი დაზიანებით, რომელიც ყველაზე მეტად ასოცირდება ინფექციურ დაავადებებთან.
- მარჩიაფავა-ბიგნამის დაავადება, დეგენერაციული დაავადება, რომელიც ხასიათდება მიელინის დაკარგვით და კორძიანი სხეულის ნეკროზით.
ისტორია რედაქტირება
პირველად კორძიანი სხეულსა და გენდერს შორის კავშირი შენიშნა რ. ბ. ბინმა, ფილადელფიელმა ანატომისტმა, რომელმაც 1906 წელს წამოაყენა მოსაზრება რომ „განსაკუთრებული ზომა კორძიანი სხეულისა შეიძლება ნიშნავდეს განსაკუთრებულ ინტელექტუალურ აქტივობას“ და რომ არსებობს გაზომვადი განსხვავებები ქალსა და კაცს შორის. შესაძლოა სწორედ იმ დროინდელი პოლიტიკური კლიმატის გავლენით, ის ამტკიცებდა კორძიანი სხეულის ზომის სხვადასხვაობას რასების სხვაობის მიხედვით. მისი მოსაზრება საბოლოოდ უარყოფილ იქნა ფრანკლინ მალის მიერ, რომელიც საკუთარ ლაბორატორიას ფლობდა.[26]
უფრო ხმაურიანი გავლენა ჰქონდა 1982 წლის ჰოლოვეისა და უტმსინგის სამეცნიერო სტატიას, რომელმაც წამოაყენა აზრი სქესობრივი განსხვავებების არსებობის შესახებ ადამიანის ტვინის მორფოლოგიაში, რომელიც მიემართებოდა განსხვავებებს კოგნიტურ უნარებში.[27] თაიმმა გამოაქვეყნა სტატია 1992 წელს, რომლითაც შემოთავაზებულ იქნა, რომ რადგანაც კორძიანი სხეული ძირითადად უფრო ფართოა ქალების ტვინებში, ვიდრე მამაკაცების ტვინებში, ეს უკანასკნელი შესაძლოა უზრუნველყოფდეს უფრო დიდ ჯვარედინ კავშირებს ნახევარსფეროებს შორის, - რაც შესაძლოა იყოს ბაზა ქალური ინტუიციისთვის.[28]
უფრო ახალმა პუბლიკაციებმა ფსიქოლოგიურ ლიტერატურაში წამოჭრა ეჭვები იმასთან დაკავშირებით მართლაც არის თუ არა კორძიანი სხეულის ზომები სხვადასხვა. 1980-იანი წლებიდან მოყოლებული, 49 კვლევის მეტა ანალიზმა, ლაკოსტე-უტამსინგის და ჰალვეის წინააღმდეგ აჩვენა, რომ არავითარი სქესობრივი განსხვავებები არ არსებობს კორძიანი სხეულის ზომებში, თანაც ისე, რომ მხედველობაში არის მიღებული მამაკაცის უფრო დიდი ტვინის მოცულობა.[26] 2006 წლის კვლევამ, რომელიც იყენებდა მრი-ს თხელ ნაჭერს, აჩვენა რომ არავითარი განსხვავებები არ არის კორძიანი სხეულის სისქეში, როდესაც გათვალისწინებულია საკვლევის სუბიექტის ზომა.[29]
სხვა ცხოველები რედაქტირება
კორძიანი სხეული აღმოჩენილია მხოლოდ პლაცენტურ ძუძუმწოვრებში, (ეუთერიანებში) კორძიანი სხეული არ გვხვდება მონოთრემებში და ჩანთოსნებში,[30] არც სხვა ხერხემლიანებში, როგორებიცაა ჩიტები, რეპტილიები, ამფიბიები და თევზები.[31] (სხვა ჯგუფებს აქვთ ტვინის სხვა სტრუქტურა, რომელიც მათ ნახევარსფეროებს შორის კავშირის საშუალებას აძლევს, როგორიც არის ანტერიორული დაკავშირება, რომელიც მსახურობს როგორც პირველდაწყებითი მეთოდი, ჩანთოსნებში ნახევარსფეროთაშორისი კომუნიკაციის უზრუნველსაყოფად)[32] [33] და ატარებს ყველა დამაკავშირებელ ბოჭკოებს, რომლებიც ამოიზრდებიან ტავის ტვინის ახალი ქერქიდან (ასევე ცნობილია როგორც ჰომოგენეტური ქერქი) მაშინ როცა, პლაცენტურ ძუძუმწოვრებში ანტერიორული დამაკავშირებელი მხოლოდ ამ ბოჭკოების რაღაც ნაწილს ატარებს.[34] პრიმატებში, ნერვების ტრანსმისიის სიჩქარე დამოკიდებულია მათი მიელინიზაციის ხარისხზე ან ცხიმოვან ფენაზე. ამაზე გავლენას ახდენს ნერვის აქსონის დიამეტრი. პრიმატების უმეტესობაში აქსონური დიამეტრი იზრდება ტვინის ზომის პირდაპირპროპორციულად, რათა დააკომპენსიროს გაზრდილი დისტანცია, იმისთვის რომ შესაძლებელი გახადოს მოგზაურობა ნერვული იმპულსების ტრანსიმისიისთვის. ეს ტვინს საშუალებას აძლევს რომ სენსორული და მოტორული იმპულსების კოორდინირება მოახდინოს. ასევე, მთლიანი ტვინის ზომის მასშტაბირება და გაზრდილი მიელინიზაცია არ გამოვლენილა შიმპანზეებში და ადამიანებში. რამაც გამოიწვია ადამიანის კორძიანი სხეულის მიერ ორმაგი დროის საჭიროება, იმისათვის რომ მოეხდინა ნახევარსფეროთაშორისი კომუნიკაცია, როგორც მაკაკებში. ბოჭკოვანი ქსელი, რომელშიც ჩნდება კორძიანი სხეული შესაძლოა გაიზარდოს ისეთ მანძილზე ადამიანებში, რომ შეიჭრას თავის ზემოთ და სოლისებრად ამაგრებდეს ჰიპოკამპურ სტრუქტურებს.[35]
გალერეა რედაქტირება
-

-
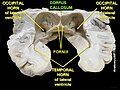 კორძიანი სხეული სხვადასხვა პროექციით
კორძიანი სხეული სხვადასხვა პროექციით -
-
 კორონალური T2. ტვინის MRI კუდიანი ბირთვის დონეზე, კორძიანი სხეულის ხაზგასმით.
კორონალური T2. ტვინის MRI კუდიანი ბირთვის დონეზე, კორძიანი სხეულის ხაზგასმით. -
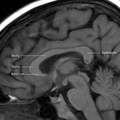 კორძიანი სხეულის ნაწილები MRI-ით.
კორძიანი სხეულის ნაწილები MRI-ით. -
DTI კორძიანი სხეული.
-
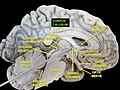
 ტვინის პარკუჭები და ბაზალური განგლია. უმაღლესი ჰორიზონტალური დათვალიერება სექციისა ღრმა გაკვეთით.
ტვინის პარკუჭები და ბაზალური განგლია. უმაღლესი ჰორიზონტალური დათვალიერება სექციისა ღრმა გაკვეთით. -

-
 თავის ტვინი. კორძიანი სხეულის ქვედა დათვალიერება ღრმა გაკვეთით.
თავის ტვინი. კორძიანი სხეულის ქვედა დათვალიერება ღრმა გაკვეთით. -










სქოლიო რედაქტირება
- ↑ 1.0 1.1 Luders, Eileen; Thompson, Paul M.; Toga, Arthur W. (18 August 2010). "The Development of the Corpus Callosum in the Healthy Human Brain". Journal of Neuroscience. pp. 10985–10990. doi:10.1523/JNEUROSCI.5122-09.2010.
- ↑ Caminiti, Roberto; Ghaziri, Hassan; Galuske, Ralf; Hof, Patrick R.; Innocenti, Giorgio M. (2009). "Evolution amplified processing with temporally dispersed slow neuronal connectivity in primates". Proceedings of the National Academy of Sciences. 106 (46): 19551–6. Bibcode:2009PNAS..10619551C. doi:10.1073/pnas.0907655106. JSTOR 25593230. PMC 2770441 . PMID 19875694.
- ↑ Hofer, Sabine; Frahm, Jens (2006). "Topography of the human corpus callosum revisited—Comprehensive fiber tractography using diffusion tensor magnetic resonance imaging". NeuroImage. 32 (3): 989–94. doi:10.1016/j.neuroimage.2006.05.044. PMID 16854598
- ↑ Davatzikos, C; Resnick, S. M. (1998). "Sex differences in anatomic measures of interhemispheric connectivity: Correlations with cognition in women but not men". Cerebral Cortex. 8 (7): 635–40. doi:10.1093/cercor/8.7.635. PMID 9823484
- ↑ Ardekani, B. A.; Figarsky, K.; Sidtis, J. J. (2012). "Sexual Dimorphism in the Human Corpus Callosum: An MRI Study Using the OASIS Brain Database". Cerebral Cortex. 23(10): 2514–20. doi:10.1093/cercor/bhs253. PMC 3767965 . PMID 22891036.
- ↑ Dubb, Abraham; Gur, Ruben; Avants, Brian; Gee, James (2003). "Characterization of sexual dimorphism in the human corpus callosum". NeuroImage. 20 (1): 512–9. doi:10.1016/S1053-8119(03)00313-6. PMID 14527611.
- ↑ Westerhausen, René; Kreuder, Frank; Sequeira, Sarah Dos Santos; Walter, Christof; Woerner, Wolfgang; Wittling, Ralf Arne; Schweiger, Elisabeth; Wittling, Werner (2004). "Effects of handedness and gender on macro- and microstructure of the corpus callosum and its subregions: A combined high-resolution and diffusion-tensor MRI study". Cognitive Brain Research. 21 (3): 418–26. doi:10.1016/j.cogbrainres.2004.07.002. PMID 15511657.
- ↑ Shin, Yong-Wook; Jin Kim, Dae; Hyon Ha, Tae; Park, Hae-Jeong; Moon, Won-Jin; Chul Chung, Eun; Min Lee, Jong; Young Kim, In; Kim, Sun I.; et al. (2005). "Sex differences in the human corpus callosum: Diffusion tensor imaging study". NeuroReport. 16 (8): 795–8. doi:10.1097/00001756-200505310-00003. PMID 15891572.
- ↑ Kontos, Despina; Megalooikonomou, Vasileios; Gee, James C. (2009). "Morphometric analysis of brain images with reduced number of statistical tests: A study on the gender-related differentiation of the corpus callosum". Artificial Intelligence in Medicine. 47 (1): 75–86. doi:10.1016/j.artmed.2009.05.007. PMC 2732126 . PMID 19559582.
- ↑ Spasojevic, Goran; Stojanovic, Zlatan; Suscevic, Dusan; Malobabic, Slobodan (2006). "Sexual dimorphism of the human corpus callosum: Digital morphometric study". Vojnosanitetski pregled. 63 (11): 933. doi:10.2298/VSP0611933S.
- ↑ Yokota, Y.; Kawamura, Y.; Kameya, Y. (2005). "Callosal Shapes at the Midsagittal Plane: MRI Differences of Normal Males, Normal Females, and GID". 2005 IEEE Engineering in Medicine and Biology 27th Annual Conference: 3055–8. doi:10.1109/IEMBS.2005.1617119. ISBN 0-7803-8741-4.
- ↑ Levitin, Daniel J. "This is Your Brain on Music", '
- ↑ 13.0 13.1 13.2 Witelson, S. (1985). "The brain connection: The corpus callosum is larger in left-handers". Science. 229 (4714): 665–8. Bibcode:1985Sci...229..665W. doi:10.1126/science.4023705. PMID 4023705.
- ↑ Driesen, Naomi R.; Raz, Naftali (1995). "The influence of sex, age, and handedness on corpus callosum morphology: A meta-analysis". Psychobiology. 23 (3): 240–7.
- ↑ Kozlovskiy, S.A.; Vartanov, A.V.; Pyasik, M.M.; Nikonova, E.Y. (2012). "Functional role of corpus callosum regions in human memory functioning". International Journal of Psychophysiology. 85 (3): 396–7. doi:10.1016/j.ijpsycho.2012.07.092.
- ↑ Hynd, G. W.; Hall, J.; Novey, E. S.; Eliopulos, D.; Black, K.; Gonzalez, J. J.; Edmonds, J. E.; Riccio, C.; Cohen, M. (1995). "Dyslexia and Corpus Callosum Morphology". Archives of Neurology. 52 (1): 32–8. doi:10.1001/archneur.1995.00540250036010. PMID 7826273.
- ↑ Von Plessen, K; Lundervold, A; Duta, N; Heiervang, E; Klauschen, F; Smievoll, AI; Ersland, L; Hugdahl, K (2002). "Less developed corpus callosum in dyslexic subjects—a structural MRI study". Neuropsychologia. 40 (7): 1035–44. doi:10.1016/S0028-3932(01)00143-9. PMID 11900755.
- ↑ Steele, C. J.; Bailey, J. A.; Zatorre, R. J.; Penhune, V. B. (2013). "Early Musical Training and White-Matter Plasticity in the Corpus Callosum: Evidence for a Sensitive Period". Journal of Neuroscience. 33 (3): 1282–90. doi:10.1523/JNEUROSCI.3578-12.2013. PMID 23325263
- ↑ Clarke, Dave F.; Wheless, James W.; Chacon, Monica M.; Breier, Joshua; Koenig, Mary-Kay; McManis, Mark; Castillo, Edward; Baumgartner, James E. (2007). "Corpus callosotomy: A palliative therapeutic technique may help identify resectable epileptogenic foci". Seizure. 16 (6): 545–53. doi:10.1016/j.seizure.2007.04.004. PMID 17521926
- ↑ "WebMd Corpus Callotomy". Web MD. July 18, 2010. Retrieved July 18, 2010.
- ↑ Rakic, P; Yakovlev, PI (January 1968). "Development of the corpus callosum and cavum septi in man". The Journal of Comparative Neurology. 132 (1): 45–72. doi:10.1002/cne.901320103. PMID 5293999
- ↑ Rash, BG; Richards, LJ (28 May 2001). "A role for cingulate pioneering axons in the development of the corpus callosum". The Journal of Comparative Neurology. 434 (2): 147–57. doi:10.1002/cne.1170. PMID 11331522
- ↑ Dobyns, W. B. (1996). "Absence makes the search grow longer". American Journal of Human Genetics. 58 (1): 7–16. PMC 1914936 . PMID 8554070
- ↑ "NINDS Agenesis of the Corpus Callosum Information Page: NINDS". RightDiagnosis.com. Retrieved Aug 30, 2011.
- ↑ "Autism May Involve A Lack Of Connections And Coordination In Separate Areas Of The Brain, Researchers Find". Medical News Today.
- ↑ 26.0 26.1 Bishop, Katherine M.; Wahlsten, Douglas (1997). "Sex Differences in the Human Corpus Callosum: Myth or Reality?". Neuroscience & Biobehavioral Reviews. 21 (5): 581–601. doi:10.1016/S0149-7634(96)00049-8. PMID 9353793.
- ↑ Delacoste-Utamsing, C; Holloway, R. (1982). "Sexual dimorphism in the human corpus callosum". Science. 216 (4553): 1431–2. Bibcode:1982Sci...216.1431D. doi:10.1126/science.7089533. PMID 7089533.
- ↑ C Gorman (20 January 1992) "Sizing up the sexes" Time: 36-43. As cited by Bishop and Wahlsten.
- ↑ Luders, Eileen; Narr, Katherine L.; Zaidel, Eran; Thompson, Paul M.; Toga, Arthur W. (2006). "Gender effects on callosal thickness in scaled and unscaled space". NeuroReport. 17 (11): 1103–6. doi:10.1097/01.wnr.0000227987.77304.cc. PMID 16837835.
- ↑ Keeler, Clyde E. (1933). "Absence of the Corpus callosum as a Mendelizing Character in the House Mouse". Proceedings of the National Academy of Sciences of the United States of America. 19 (6): 609–11. Bibcode:1933PNAS...19..609K.
- ↑ Sarnat, Harvey B., and Paolo Curatolo (2007). Malformations of the Nervous System: Handbook of Clinical Neurology, p. 68 დაარქივებული 2015-11-26 საიტზე Wayback Machine.
- ↑ Ashwell, Ken (2010). The Neurobiology of Australian Marsupials: Brain Evolution in the Other Mammalian Radiation, p. 50
- ↑ Armati, Patricia J., Chris R. Dickman, and Ian D. Hume (2006). Marsupials, p. 175
- ↑ Butler, Ann B., and William Hodos (2005). Comparative Vertebrate Neuroanatomy: Evolution and Adaptation, p. 361
- ↑ Morris, H., & Schaeffer, J. P. (1953). The Nervous system-The Brain or Encephalon. Human anatomy; a complete systematic treatise. (11th ed., pp. 920–921, 964–965). New York: Blakiston.